Hay potenciales eléctricos en todas las membranas de todas las células del cuerpo; algunas células como las nerviosas y las musculares, son excitables, es decir capaces de auto generar impulsos electroquímicos en sus membranas. En mayor parte de los casos estos impulsos sirven para transmitir señales a lo largo de la membrana. En otros tipos de células, como las glandulares, macrófagos y células ciliadas, es probable que ocurran alteraciones de otro tipo en el potencial de la membrana y esos cambios desempeñan una función significativa en el control de muchas funciones celulares. La existencia de una diferencia de potencial es ante todo una indicación de la existencia de una separación de cargas. De alguna manera las membranas biológicas contribuyen a que se mantenga un exceso relativo de cargas negativas en el interior celular con respecto al medio extracelular.
En principio podríamos distinguir varios motivos por los que existe una separación de cargas a través de diversos tipos de membranas biológicas. Por un lado, muchos de los componentes estructurales de la membrana son ionizables a pH celular y podría pensarse que una distribución asimétrica de estos grupos con diferentes pK daría lugar a una diferencia de potencial similar a la observada. Por otro lado, algunas proteínas transportadoras acopladas a la hidrólisis de ATP, notablemente la bomba de sodio y potasio, son electrogénicas. Esto significa que en la operación normal del transportador se separan cargas, o dicho de otra forma, son generadores de corriente. Por último, en situaciones en las que existen gradientes de potencial electroquímico de iones a través de la membrana pueden aparecer potenciales de difusión.
Con respecto a los componentes cargados de las membranas biológicas, debe tenerse en cuenta que la presencia de un electrolito en un medio en el que las cargas pueden moverse libremente provocaría la neutralización parcial de la carga del mismo, por parte de los contraiones en solución. Sin embargo no es imposible que de existir diferencias de densidad superficial de carga entre ambas láminas de las bicapas, éstas contribuyan a la generación o estabilización del potencial de membrana. Se ha propuesto que estas cargas, si bien no contribuyen al potencial transmembrana global, pueden tener un efecto local sobre las proteínas de transporte.
Cuando la concentración de potasio es muy alta dentro de la célula y muy baja fuera de ella a esto se le llama permeabilidad selectiva los iones de potasio pero a ningunos más. A causa del enorme gradiente de concentración entre el potasio interior y el exterior, los iones de potasio muestran fuerte tendencia a difundirse hacia fuera. Al difundirse se llevan consigo cargas positivas hacia el exterior generando un estado de electropositividad fuera de la membrana y de electronegatividad en el interior debido a los aniones negativos que no se difunden al exterior junto con el potasio. Esta nueva diferencia de potencial rechaza los iones positivos de potasio en dirección retrograda desde el exterior hacia el interior.
Cuando hay una concentración muy baja de iones de sodio fuera de la membrana y una concertación muy baja de sodio en el interior. Estos iones también tienen carga positiva y la membrana es muy permeable al sodio e impermeable a otros iones. La difusión de los iones de sodio hacia el interior genera un potencial de membrana ahora de polaridad opuesta; el lado externo es negativo y el lado interno es positivo. Una ves mas los milisegundos el potencial de membrana se eleva lo suficiente para bloquear la difusión neta de iones de sodio hacia el interior; sin embargo, en esta ocasión el potencial delos troncos nerviosos de mamíferos se aproxima a 61mV y el interior de la fibra es positivo.
Esta es la diferencia de concentración de iones a través de una membrana con permeabilidad selectiva puede generar un potencial de membrana en condiciones apropiadas.
Si colocamos entre dos compartimentos una membrana biológica, la cual sea permeable sólo a los iones K y se agrega una solución de KCl en uno de los compartimentos, los iones K comenzarán a moverse siguiendo su gradiente de concentración.
Como los iones Cl no pueden atravesar la membrana, ésta se carga negativamente de ese lado y positivamente del otro.
Como los iones potasio al quedar del lado positivo se sienten rechazados por esta carga tender n a regresar al compartimiento inicial, se dice que se mueve en contra de su gradiente eléctrico. Por lo tanto:
- El movimiento de los iones es en favor de su gradiente químico y en contra del eléctrico.
- El movimiento de los iones K continuar hasta equilibrar estas dos fuerzas.
- Es decir hasta alcanzar el potencial de equilibrio electroquímico para el ion potasio.
Si se determina la diferencia de potencial entre estos compartimentos tendrá un valor de -90 mV Si la membrana celular fuera permeable a un solo ion el potencial de membrana en reposo seria igual al potencial de equilibrio para ese ion. Como la membrana es permeable en diferente grado a los iones K, Na y Cl, el valor del potencial se calcula combinando las cifras de los potenciales de equilibrio para dichos iones de acuerdo con la permeabilidad de la membrana a cada uno. Así el K y el Cl para los cuales la membrana es más permeable, contribuyen m s al potencial de membrana que el Na. Es importante saber que en reposo la membrana es 100 veces más permeable al K y al Cl que al Na.
Otro factor que participa en el mantenimiento del potencial de membrana en reposo es la presencia de bombas metabólicas. Una de las bombas que mantiene el gradiente es la de a/K. Bombas como éstas mantienen una diferencia de concentraciones de los iones porque son electrógenas, ya que sacan tres iones sodio e introducen al citoplasma dos iones K.
Él ultimo factor que participa en el potencial de membrana en reposo, pero no por eso el menos importante, es la presencia de proteínas en el citoplasma. Las cuales por ser aniones tan grandes no atraviesan la membrana y contribuyen a mantener la carga negativa del interior.
También es importante hablar de los potenciales electroquímicos debido a que éstos son los responsables directos que casi todos lo fenómenos eléctricos que tienen lugar en el cuerpo de los animales.
Una neurona está polarizada, es decir, tiene una carga eléctrica negativa en el interior de la membrana celular respecto al exterior. Esto se debe a la libre circulación de iones potasio con carga positiva a través de la membrana celular, y al mismo tiempo, a la retención de moléculas grandes con carga negativa dentro de la célula. Los iones de sodio con carga positiva se mantienen en el exterior de la célula mediante un proceso activo. Todas las células tienen esta diferencia de potencial, pero cuando se aplica a una célula nerviosa una corriente estimuladora se produce un suceso único. Primero, los iones de potasio penetran en la célula, reduciendo su carga negativa despolarización. En un cierto momento las propiedades de la membrana cambian y la célula se hace permeable al sodio, que entra en ella con rapidez y origina una carga neta positiva en el interior de la neurona. Esto se denomina el potencial de acción.
Una vez alcanzado este potencial en una zona de la neurona, éste se propaga a lo largo del axón mediante un intercambio de iones en unos puntos específicos llamados nódulos de Ranvier. La amplitud del potencial de acción es autolimitado, debido a que una concentración elevada de sodio en el interior origina la expulsión de la célula primero de iones potasio, y después de sodio, restableciendo la carga negativa en el interior de la membrana celular, es decir la neurona sé repolariza. El proceso completo dura menos de una milésima de segundo. Después de un breve lapso, llamado periodo refractario, la neurona está en condiciones de repetir este proceso.
La contracción sincronizada de todas las células que están acopladas eléctricamente constituyendo el tejido cardíaco, generan la contracción sincrónica de cada una de las cámaras del corazón. La contracción de cada célula está asociada a un potencial de acción.
En es estado de reposo, antes iniciarse el potencial de acción, la conductancia para los iones potasio es de 50 a 100 veces mayor que la escape mucho mayor de iones sodio. Esto se debe al escape mucho mayor de iones de potasio en comparación con el de iones de sodio a través de los canales de escape.
El inicio del potencial de acción también abre los canales del potasio mediante la compuerta de voltaje; esta apertura empieza una fraccion de milisegundos después de abrirse los canales del sodio. Al final del potencial de acción, el potencial de membrana retorna a su estado negativo y los canales del potasio se cierran de nuevo regresando a su estado original, pero una vez mas solo después de un pequeño retraso.
Justo al inicio del potencial de acción, el valor de esta relación aumenta mas de 1000 veces. Por tanto, los iones sodio penetran a la fibra en cantidad mucho mayor que la cantidad de iones de potasio que sale. Esta es la razón de que el potencial de membrana se haga positivo. Los canales del sodio comienzan a inactivarse y al mismo tiempo se abren los canales del potasio de modo que ahora la relación de conductancia se modifica a favor de un aumento en la conductancia al potasio y una reducción de la conductancia del sodio. Esto permite una salida muy rápida de iones potasio, en tanto la entrada de sodio a la fibra es prácticamente nula. Por consiguiente, el potencial de acción retorna de inmediato al nivel basal.
No habrá potencial que desencadene el potencial de acción en tanto la elevación inicial del potencial de membrana no sea lo bastante grande para crear el circulo. Por lo general se requiere una elevación súbita de 15 a 30 mV en el potencial de membrana. Por tanto, al incrementar de modo repentino el potencial de membrana, de -90mV a cerca de -64 mV, en una fibra gruesa casi siempre se genera un potencial de acción de manera súbita. A este nivel de -65mV se le denomina umbral de estimulación.
Una corriente procedente de un electrodo, o en el caso que nos ocupa, la onda de excitación procedente del marcapasos, añade cargas positivas al lado intracelular de la membrana reduciendo el potencial de reposo y provocando una lenta despolarización de la membrana. Este comportamiento se representa desde un punto a a un punto b.
A medida que el potencial de la membrana V se aproxima al umbral Vu, se abren los canales de iones Na en la membrana permitiendo su paso al interior de la misma. De este modo se consigue un equilibrio entre los iones Na que entran y los iones K que salen para compensar la entrada de carga positiva en el interior provocada por los iones Na. Este proceso tiene lugar de esta manera debido a la existencia de un potencial en la membrana y debido a que hay una mayor concentración de iones Na y K en el exterior y en el interior de la membrana respectivamente. Este proceso no se realiza indefinidamente sino que se alcanza un equilibrio cuando la diferencia de potencial debida al gradiente de concentración es igual al de repulsión debido a la carga a ambos lados de la membrana. Cuando V=Vu entonces los iones Na exceden a los K, esto ocurre en el punto b.
Esta entrada neta de carga, hace que la membrana se despolarice más. Esta despolarización es regenerada ya que a medida que aumenta la carga aumenta el potencial positivo lo que hacen que se abran nuevos canales para el Na. Esto produce la rápida subida del PA.
A medida que el potencial de membrana Vm se aproxima al potencial de equilibrio de los iones Na la entrada de iones Na a la célula se hace progresivamente menor haciendo que la tasa de cambio de potencial de haga más lenta. Esto se observa desde el punto c al d.
Ahora los canales de iones Na abiertos se inactivan haciendo que el PA disminuya gradualmente restableciendo el potencial de reposo. A este proceso se le denomina proceso de repolarización.
El proceso de repolarización se acelera por la apertura de los canales K dependientes del voltaje. Este flujo de salida de iones K elimina carga positiva de la célula.
En una membrana excitable no hay una dirección definida para que el impulso se propague, si no que el potencial de acción puede viajar en ambas direcciones alejándose del estimulo por todas las ramificaciones de una fibra nerviosa hasta despolarizar toda la membrana en su conjunto.
Una ves iniciado un potencial de acción en cualquier punto de la membrana de una fibra normal, el proceso de despolarización de desplaza sobre toda la membrana si las condiciones son adecuadas o no se desplaza nada en condiciones inadecuadas. A ello se le denomina respuesta del todo o nada.
Las fibras nerviosas están constituidas por un axón rodeado (fibras mielínicas, de conducción rápida) o no (fibras amielínicas, de conducción lenta) de múltiples capas de membrana celular (mielina) de una célula de Schwann, que se enrolla alrededor del axón. Esta capa mielínica está interrumpida periódicamente en los nódulos de Ranvier, puntos `saltatorios' de los impulsos eléctricos.
Un tronco nervioso típico contiene unas pocas fibras nerviosas gruesas que ocupan la mayor parte del área de sección transversal y un numero mucho mayor de fibras delgadas situadas entre las gruesas. Las fibras gruesas son mielinicas y las delgadas amielinicas. El tronco nervioso de grosor promedio contiene casi el doble de fibras amielinicas que mielinicas.
El cordón central de la fibra es el axon y la membrana de axon es la verdadera membrana conductora. El centro del axon es la verdadera membrana conductora. El centro del axon esta lleno de citoplasma, un liquido intracelular viscoso. Rodeando el axon se encuentra una vaina de mielina que suele ser mas gruesa que el propio axon; los nodos de Ranvier interrumpen la capa de mielina de toda su extensión de intervalos de1 a 3 milimetros.
Las células de Schwann forman capas de mielina que rodean el axon de la manera siguiente: primero, la membrana de una celula de Swchann envuelve el axon, luego la célula gira muchas veces alrededor del axon dejando tras de sí muchas capas de membrana elular que contienen una sustancia lipida denominada esfingomielina. Esta sustancia es un excelente aislante que reduce el flujo de ionice a través de la membrana casi 5000 veces.
Conducción saltatoria de nodo en las fibras mielinizadas.
Aunque no haya flujo significativo de iones a través de las espesas capas de mielina de las fibras milinizadas los iones aun pueden fluir con gran facilidad a través de los nodos de Ranvier. Por tanto lo potenciales de acción pueden ocurrir solo en los nodos. Así , el potencial de acción se conduce de un nodo a otro, este fenómeno se denomina conducción saltatoria. Esto es, la corriente eléctrica fluye de un nodo a través del liquido extracelular circunvecino y del axoplasma excitando consecutivamente nodos sucesivos. El impulso nervioso salta a lo largo de la fibra; este es el origen del nombre saltatorio.
Básicamente todo factor que inicie l a difusión de iones de sodio al interior de la célula a traves de la membrana en cantidad sificiente puede desencadenar la abertura automatica y regenrativa de los canales del sodio.
Todos lo medio se utilizan en diferentes puntos del cuerpo para despertar potenciales de accion en nervio o en músculo: preción mecanica para excitar terminaciones nerviosas sensoriales en a piel, neurotransmisores químicos para transmitir señales de una neurona a la siguiente en el cerebro y corriente electrica para transmitir señales entre las fibras musculares del corazón y del intestino.
La causa de este efecto es la siguiente: los potenciales de acción se inician por abertura de la compuerta de voltaje de los canales del sodio.
La corriente negativa del electrodo negativo reduce de inmediato el voltaje fuera de la membrana aproximándolo al potencial negativo del interior de la fibra.
Al reducirse el voltaje electrico a través de la membrana, los canales del sodio se activan y se produce un potencial de acción.
Un estímulo es la forma de interacción entre el ser vivo y el medio, es el agente, condición o energía capaz de provocar una respuesta en un organismo determinado.
Los distintos estimulos que originan la despolarizacion celular se han clasificado entres tipos distintos: químicos, mecánicos y eléctricos.
Se considera un estimulo químico a cualquier sustancia química que, tras la union a un repector situado en la membrana de la neurona, determina la apertura o cierre de canales ionicos, lo cual hara que se establezca la apertura automatica de los canales de sodio que dan a lugar a la aparición de un potencial de accion. Entre estas sustancias encontramos: acidos, bases, soluciones salinas de elevada concentración etc.
Son estimulos mecánicos aquellos que causan las alteaciones en la energia mecanica de la neurona ( como vibración, pinchazo, etc), lo que implica una brusca penetración de sodio que se desencadenara el potencial de accion.
Son estimulos eléctricos los que que cambias la carga electrica de las neuronas, añadiendo cargas positivas o negativas. La corriente electrica inducida de manera artificial mediante la implantación de un par de electrodos intra y extracelulares, conectados a un generador de corriente.




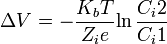


{kind=link}